Learning Objectives
After reading this chapter, students should be able to:

discuss the general origins and spread of anatomically modern humans
trace the development of
H. neanderthalensis in Eurasia
trace the development of
H. sapiens in Africa
describe the major sites and archaeological cultures providing evidence for this period
understand the arguments for and against the replacement vs. the multiregional origin models for the origins of modern humans
explain the profound behavioral changes associated with modern humans, such as new technology, and the use of symbolism and art
understand the difficulties of dating and interpreting fossils during this transition
describe the role of DNA research in the overall picture of Middle and Upper Paleolithic studies
discuss the debate over the relationship between modern humans and Neanderthals
trace the causes of differing biological and cultural evolution among Neanderthals and modern humans
explain current theories about the colonization of Europe, Asia, Australia, and the Americas
THE CLIMATIC BACKGROUND
Modern humans evolved in a context of global climatic fluctuation, reflected in oxygen isotope ratios in ice and deep-sea cores. Over the last 400,000 years, there were at least four interglacials, four severe glacials, and stadials/interstadials within and between these. Geological and faunal indicators are correlated to these.
In the north, plant communities flourished or died back, in Africa, wetter or arid periods alternated, allowing human populations to spread initially, but when desert conditions returned, small groups of humans may have been forced to migrate north or east out of Africa. In eastern Asia, climate was relatively stable, perhaps explaining stasis among
Homo erectus.
COMPETING HYPOTHESES FOR THE ORIGIN OF HOMO SAPIENS
In the 1940s, a multi-regional origin model was first proposed:
Homo sapiens arose from
Homo ergaster populations in different areas after they moved outside of Africa. Gene flow between populations at the edges of larger regions prevented divergence into more then one species. This "multi-regional evolution hypothesis" has several current proponents.
By the 1960s, others suggested that modern humans originated in Africa, and then moved out already evolved. This theory has been called Noah's Ark, Garden of Eden, single origins, and Out of Africa hypotheses.
Proponents of these theories never regarded them as mutually exclusive, but they have had a polarizing effect. Intermediary models have also been debated since the late 1980s. Modern genetics and ancient DNA have provided strong but not unequivocal support for a single African origin.
The Multi-regional Evolution Hypothesis In this model, modern "racial" differences have deep evolutionary roots, although gene flow maintains reproductive and anatomical similarity. Fossils from Asia and Australia are interpreted as showing regional continuity. The hypothesis predicts that modern human fossils should appear at broadly the same time throughout the Old World; that transitional fossils between
Homo ergaster or
H. heidelbergensis and modern humans should be found; and that continuity should also occur. Genetically, modern human genes should have a deep ancestry, and genetic variability should be similar across the Old World, reflecting similar rates of change.
The Out of Africa Hypothesis This hypothesis assumes that modern humans dispersed from a single center of origin, but also that they completely replaced older, indigenous forms of
Homo, with some possibile interbreeding. The hypothesis predicts that the earliest transitional and then modern human fossils will be found only in Africa. It also suggests that modern-day humans outside of Africa will not share heritage with earlier populations in the same region. Only in Africa will great genetic diversity be found, reflecting a longer period of random mutation.
Other Hypotheses and Attempts at Consensus In the 1980s, a "hybridization and replacement model" that attempted a synthesis was proposed: regional continuity combined with slow expansions out of Africa of emerging modern humans, and varying degrees of genetic mixing. A related assimilation hypothesis was soon after proposed, in which an emergent African
Homo sapiens assimilated regional early humans into the modern human gene pool. Recently, some have suggested that multiple dispersals out of Africa were most probable, which is supported by recent genetic work.
Thus, the four main hypotheses are: 1) an African replacement hypothesis; 2) an African hybridization and replacement hypothesis; 3) an assimilation hypothesis; and 4) a multi-regional evolution hypothesis.
Fossils and genetics may fit more than one hypothesis if they are poorly defined, so their predictions bear examination: replacement and hybridization hypotheses both predict a relatively recent African origin for modern humans, and a high level of replacement during their dispersal(s) out of Africa. The extent of gene flow between
Homo sapiens and regional archaic
Homo species distinguishes them, with replacement having less gene flow and hybridization having more. In contrast, the multi-regional hypothesis assumes that the evolutionary origin in more than one region, and a much longer chronological period of emergence. The assimilation hypothesis does not necessitate such a long chronology.
Scholarly opinion is moving toward a consensus that modern humans evolved biologically in Africa. Even the complex Asian fossil record that is used to support multi-regionalism, no longer refutes African origins.
Research now centers on how many dispersal events occurred, how these are reflected in modern human genetics; whether language origin was crucial to the process; and whether modern behavior emerged gradually or suddenly. The subsequent spread of modern humans to Australia and the Americas is still hotly contested, and whether own species exchanged ideas, genes, or violence with archaic
Homo species.
THE ANATOMY OF HOMO SAPIENS
Characteristics of modern Homo include cranial capacity greater than 1350 cc; a vertical frontal bone; a high and parallel-walled cranial vault; a rounded occipital region lacking a prominent occipital torus and with a relatively flat angle of the cranial base; a non-continuous brow ridge expressed more clearly in males; a flat, non-projecting face below the expanded frontal braincase region; and a distinct chin. Postcranially, modern
Homo sapiens have long limbs with long distal limb segments, which are less robust than archaic
Homo species. A tall, long-limbed, short-trunked body, adapted to hot climates, supports an African origin.
EVOLUTION IN LOW LATITUDES: EVIDENCE FOR THE RISE OF MODERN HUMANS IN AFRICA
Fossils pertinent to an African origin are few. Three chronological groupings are: 1) earlier than 250,000 years ago, in which
Homo sapiens traits are evident but retentions from
Homo ergaster are also evident; 2) between 250,000 and 125,000 years ago, a transitional group with variable primitive and modern traits; and 3) less than 125,000 years ago, clear anatomically modern humans. A degree of morphological continuity is observable between groups.
Earliest Homo sapiens The sites include Bodo, Broken Hill, Elandsfontein (Saldanha), and Ndutu, with fossils classified variously as
Homo heidelbergensis, H. leakeyi, H. rhodesiensis, and archaic
H. sapiens.
Transitional Homo sapiens The second group come from the sites of Florisbad, Omo (Kibish), and Jebel Irhoud, with specimens variously classified as H. helmei and H. sapiens.
Anatomically Modern Humans Fossils dating after 125,000 years ago clearly belong to modern
Homo sapiens. Important sites include: Die Kelders Cave, Equus Cave, the Cave of Hearths at Makapansgat, Border Cave, Klasies River Mouth, Dar-es-Soltan, Taforalt, Afalou, Nazlet Khater, Taramsa, Mugharet es-Skhul, and Jebel Qafzeh
GENETIC KEYS TO THE ORIGINS OF MODERN HUMANS
Beginning in the 1980s, different measures of genetic variability were tested, especially in the work of Rebecca Cann, including blood groups, mitochondrial DNA, and the Y chromosome, which support a single origin. Interpretations were debated through the 1990s.
Mitochondrial DNA and the Theory of an Early African "Coalescence" Cells contain two genomes: nuclear DNA and mitochondrial DNA (mtDNA), which resides not in the cell's nucleus but in its walls. MtDNA is ideal for human evolutionary research, because it is not influenced by natural selection. As it is only inherited through the female line, variability cannot be introduced by recombination with a father's mtDNA. Only random mutation is responsible for variation in mtDNA.
As many as 10 random mtDNA mutations occur per 1000 reproductions, so a rough mutation rate or "molecular clock" can provide estimated dates for when a set of genes in a group first appeared, known as coalescence. Variability can be analyzed statistically and a phylogeny can be drawn up. Variability of living human mtDNA is surprisingly low, suggesting very recent coalescence or a population bottleneck.
When published, this data was interpreted in favor of both hypotheses, as both assume some degree of evolution in Africa. The timing of coalescence is crucial to support one theory over the other. Multi-regional hypotheses predict coalescence about 2 million years ago, while the single origins hypotheses predict it around 200,000 years ago.
After much research, estimates of
Homo sapiens coalescence all fall around 200,000 years ago (mtDNA), a little earlier (nuclear DNA), or a little later (Y chromosome), with a range of 270,000 to 200,000 years ago. Genetic diversity is greatest in sub-Saharan Africa, also supporting a single origin place.
Other Theories and Potential Consensus Mutation rates must be estimated properly for these "clocks" to be reliable. Such estimates suggest that divergence was about 2-4 percent per million years. Some have suggested that if mutation was slower, the date of coalescence would be older, perhaps supporting the multi-regional hypothesis. Misconceptions include the idea that all modern humans can be traced back to one female, "mitochondrial Eve." The mitochondrial ancestor was not the only female living, or the only one who passed on mtDNA, but part of a population.
These problems have now been addressed, providing a secure basis for estimating coalescence as late as around 150,000 years ago. As coalescence relates to specific genes, not a whole population, there are still some questions. Most genetic evidence is consistent with an African origin, but might include significant genetic contributions from Asia. High genetic variability in sub-Saharan African still points to primacy. Emerging consensus is that hybridization and replacement is best supported by the data, that long-term population size in Africa was largest, and that genes flowed out of Africa more frequently than into it. Low levels of mtDNA diversity may indicate a population bottleneck, when as few as 1500 female humans lived, perhaps due to environmental conditions, or our population size may always have been small.
Mitochondrial DNA and the Evolution of Homo neanderthalensis MtDNA from Neanderthals was sequenced in 1997, and shows genetic dissimilarity between Neanderthals and modern humans, although whether they could interbreed and produce fertile offspring or not is unknown. The differences between the species suggest half a million years of evolutionary divergence and that Neanderthals did not contribute mtDNA to the contemporary human gene pool.
ARCHAEOLOGY AND THE EMERGENCE OF "MODERN" BEHAVIOR IN MIDDLE STONE AGE AFRICA
After 100,000 years ago, new behaviors appeared, related to the biological emergence of anatomically modern
Homo sapiens and a new cognitive threshold. Once believed to have occurred later, in Europe, we now know that African Middle Stone Age people displayed modern behavior by 150,000 years ago: personal ornamentation, use of pigments, and possible mortuary rituals.
Artifactual Evidence Modern behavior in the African Middle Stone Age includes bone points, geometric microliths, and composite tools.
Hunting and Dietary Evidence Modern broad-spectrum diet based on many varied rather than few large species, more plants, and a new incorporation of marine resources appears in Africa by c. 130,000-115,000.
Evidence of Site Modification and Art Modern behavior includes organized space, including dwellings, and is rare in Neanderthal contexts, and also in the African Middle Stone Age.
Blombos Cave (South Africa) exemplifies most aspects of a modern way of life. Modernity, according to current data, had a late and relatively sudden emergence rather than a slow development.
EVOLUTION IN HIGH LATITUDES: THE NEANDERTHALS
African
Homo sapiens differed most drastically in cranial shape with little facial change, while European Neanderthals retained primitive crania but underwent facial modifications.
The Anatomy of Homo neanderthalensis Neanderthal traits began to appear 450,000 years ago and coalesced by 250,000 years ago, coinciding with increased glacial cycles, and are most pronounced in northern latitudes, suggesting biological adaptation to cold, dry environments and a physically demanding lifestyle.
Neanderthals retained cranial vaults like
H. heidelbergensis and possessed powerful biting musculature. Wear on front teeth suggests use as a vice-like tool. The nasal aperture and sinuses were large, and probably warmed and moistened cold, dry tundra air to protect the brain and respiration. The brain was generally larger than H. sapiens and postcranially, a large body with robust limbs and pronounced muscular insertions were adaptations to habitual physical stress. Relatively short limbs retained body heat.
Exploitation of Resources: Hunting, Gathering, and Scavenging Most Neanderthals died by their early 40s, a few lived into their 50s. Mortality peaked in infancy and adolescence, coinciding with weaning, then adult activities. Numerous healed upper body fractures closely parallel modern injuries among rodeo riders, suggesting close-range hunting.
Charred plant remains at some sites indicate that they were important in the diet when available, although stable isotope analyses of bone suggest a diet of up to 90 percent meat. Large, medium, and small animals were scavenged and hunted with hand-thrust spears of wood or with stone points.
The lack of a dominant prey species indicates opportunistic hunting strategies, with occasional planned cliff-drives of mammoth and woolly rhinoceros as early as 200,000 years ago. After 150,000 years ago, planning is suggested through targeting of adult bison and aurochs. Neanderthals probably employed many hunting methods.
The Mousterian Lithic Industry Neanderthals produced a Middle Paleolithic lithic industry called Mousterian (after Le Moustier in France). They used the Levallois flaking technique to produce predictably sized flakes and blades, and the discoidal method, yielding as many variously sized flakes as possible. Some were unmodified, others retouched into scrapers, denticulates, notches, points, and bifacial hand axes.
François Bordes suggested that five toolkit variations indicated different Neanderthal groups. This was challenged by Lewis Binford, who suggested they reflected assemblages for different tasks. Paul Mellars noted chronological patterning, with some earlier and others later, also contradicting Bordes. Lack of correlation between toolkits and differing environments or activities undermines Binford's theory. This ongoing debate is known as the "Mousterian debate." What can be said is that Neanderthal technology was flexible depending on available material.
Neanderthal Behavior Neanderthals also worked wood, antler, and bone, studied through pseudomorphs of items such as wooden posts and spears. They commonly constructed simple hearths for warmth, light, and cooking using wood or coal, and sometimes paved wet floors with stones. They used certain areas for trash disposal, toolmaking, eating, resting. Some groups buried some of their dead -- at least 20 clear examples are known, some with multiple burials over long periods. Ocher fragments and shells are found at several sites, but there is no convincing art.
EARLY DISPERSALS OF HOMO SAPIENS INTO THE LEVANTINE CORRIDOR
The Levant remained a major corridor for movement between Africa and Eurasia. Data suggest that Neanderthals and modern humans were, on occasion, contemporary in this region. Modern humans appeared by 110,000-90,000 years ago at caves in Israel, with possible "grave goods" indicating mortuary rituals, while Neanderthals buried at nearby caves between 60,000 and 50,000 years ago had no ritual indications. Neanderthal burials occur as late as 45,000 years ago but may be as old as 120,000 years ago.
The first unequivocal modern human burials with Upper Paleolithic artifacts occur around 35,000 years ago, although Aurignacian stone and bone tools and bone jewelry from 45,000 years ago indicate they may have been in the Levant earlier.
It may be that several discrete occupations reflect major biogeographic changes, with Neanderthals in the region during the severe cold conditions between about 71,000 and 60,000 years ago, when the earlier modern humans had become locally extinct.
Archaeological evidence indicates that at 50,000 years ago, both populations employed Middle Paleolithic Levalloiso-Mousterian toolkits.
However, seasonal exploitation and mobility strategies differed. Levantine Neanderthals used large, multi-purpose sites to organize activities carried out at specialized peripheral sites, and were highly predatory within a small area, hunting throughout the year. Modern humans moved between several seasonal camps located in different environments, where different activities were practiced. This required less weapon production and incorporated gathering: more energy and nutritionally efficient. Neanderthals may have ultimately been out-paced by modern humans.
THE COLONIZATION OF EAST ASIA AND AUSTRALIA
By 40,000 years ago, modern humans had dispersed into South Asia and Australia, the Caucasus and southeast Siberia, and Europe. New technological and social behaviors in the Upper Paleolithic may be linked to modern human expansion.
By c. 40,000 years ago
Homo sapiens were present in Sundaland, which was always separated from Sahul Land (Australia and New Guinea) by deep waters, indicating that ocean crossings were necessary. The actual date of colonization has long been debated. Radiocarbon dates cluster around 40,000 years ago, but many believe colonization occurred much earlier, based partly on erroneous, now corrected radiocarbon dates. Some humans may have reached the continent earlier, but an occupational watershed occurred at 40,000 years ago. Some scholars believe the fossil record shows colonization by two biologically different human populations, one from Javanese Homo erectus, and a later gracile wave from China. In Australia, there are clearly no taxonomic differences between human fossils.
THE COLONIZATION OF EUROPE, AND THE MIDDLE TO UPPER PALEOLITHIC TRANSITION
The distinctive earliest Upper Paleolithic industry in Europe, the Aurignacian (after Aurignac rockshelter, France), indicates that
Homo sapiens reached southern Europe by around 45,000 years ago, and expanded along the coastal Mediterranean as far as Iberia by 40,000 years ago, due to mild climatic conditions, seen at
Bacho Kiro and Temnata (Bulgaria)
Grotta de Fumane (Italy)
El Castillo (Spain)
General consensus is that the Aurignacian was produced only by
Homo sapiens and not by indigenous Neanderthals.
The Aurignacian Aurignacian toolkits are defined by carinated and nosed-end scrapers, retouched, "strangulated" blades, small, retouched bladelets, and various bone and antler points. Aurignacian tools are generalized and functionally flexible, important for a pioneer population, and more efficient than Neanderthal tools.
Aurignacians were well established in Europe by 35,000 years ago and on the northern European plain by around 30,000 years ago. From this time, jewelry from teeth, ivory and shell circulated over a large, established exchange network. Prismatic blade technology was widely employed, and bone and antler points, awls, and needles are related to sophisticated weaponry and elaborate tailored clothing, aiding expansion into Siberia. Art appeared: engraved ivory and bone, and painted and engraved cave walls. Examples include:
Hohle Fels cave (Germany)
Vogelherd cave (Germany)
Geissenklösterle cave (Germany)
Chauvet cave (France)
There are no convincing burials or dwelling structures, suggesting that a behavioral revolution was incomplete.
Neanderthal "Transitional" Industries The Aurignacian overlaps in some regions with late Neanderthals, who used late Middle Paleolithic or "transitional" industries until 30,000 - 28,000 years ago. A period of co-existence and interaction probably occurred. It is assumed that the Aurignacians' hunting abilities, broader spectrum economies, language, symbolism, and art created advantages over Neanderthals.
Transitional Neanderthal industries are largely Middle Paleolithic with added blade technology and Upper Paleolithic tools. They are restricted to circumscribed regions of Europe, and suggest that late Neanderthals had improved their hunting.
Relations between Neanderthals and Incoming Homo sapiens? The mix of Middle and Upper Paleolithic elements in transitional tool assemblages and ornamentation has been characterized as acculturation of Neanderthals by incoming modern humans. Other evidence is predicated upon fossils with evidence of interbreeding, although this is disputed. Sites with specific evidence include:
Grotte du Renne (France)
Pestera cu Oase (Romania)
Lagar Velho (Portugal)
Paul Mellars notes that chronologically, the overlap seems non-existent or brief in some areas, while elsewhere it may have lasted for 10,000 years. Probably more than one cause accounts for the disappearance of Neanderthals.
DEVELOPMENTS IN MODERN BEHAVIOR: THE EUROPEAN UPPER PALEOLITHIC
The Gravettian
The Aurignacian ended around 28,000 years ago, and in central Europe, out of it developed the Gravettian technocomplex (after La Gravette, France): more regionally distinct groups sharing general characteristics over a large area, between 29,000 to 21,000 years ago (the middle Upper Paleolithic), and as late as c. 14,000 years ago in southern and eastern Europe, referred to as Epigravettian. Behavioral innovations mark the period, but were ended by the Last Glacial Maximum. These include semi-sedentism, burial, and projectile technology. Technologically, it is characterized by
prismatic blade technology and a variety of points with steep backing to facilitate hafting
burins and end-scrapers for working of skins, wood, bone, and antler
small (possible bow and arrow) points that divide the period into chronological phases.
javelins thrown with a spearthrower or "atlatl"
Gravettian Behavior
A number of regional centers of occupation are recognized in the middle Upper Paleolithic -- southwest France, central Europe, Moravia (Czech Republic) and Slovakia. Central European sites were occupied for many months and much effort was invested in dwellings. Pavlov and Dolní Vestonice I and II in Moravia have yielded
hundreds of bones of mammoth, bison, and reindeer
complex patterning suggestive of highly organized campsites
simple firing of loess into small animal figurines, perhaps as part of a ritual
impressions of sophisticated cordage, basketry, and textiles as early as 27,000 years ago
Sites on the River Don in Russia, such as Kostenki I, yielded semi-subterranean dwellings, roofed with mammoth ribs and furs, organized in an oval around nine hearths. Other features include
pits cut into the permafrost containing large herbivore bones, indicating meat storage
bones of fish and small animals suggesting a broad spectrum diet
stable isotope analysis indicates that riverine resources contributed up to 50 percent of protein.
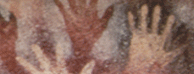
Over 40 burials, some double and two triple, with a high degree of ornamentation and grave offerings, attest to the development of mortuary activity, such as at Sungir' (Russia). Symbolic and artistic behavior is seen in "Venus" figurines and continued cave painting and engraving, with hand prints and stencils, human figures and herbivores depicted on complex, multi-phased panels Abundant art may reflect the social importance of symbolic activity during the climatic deterioration approaching the Last Glacial Maximum.
The Magdelenian
The climate grew most severe between 21,000 and 18,000 years ago, and northern and high-altitude areas were abandoned. Humans crowded into southern European refuges, undergoing marked regionalization due to isolation. In Italy, the Epigravettian, shows clear continuity, but in France and Iberia, new cultures emerged, such as the Solutrean (after Solutré, France).
Beginning 18,000 years ago, climate improved and Europe was rapidly recolonized. Late Upper Paleolithic culture is referred to as the Magdalenian technocomplex (after La Madeleine rockshelter, France). First seen in Iberia and southern France, the culture spread to Russia and southern Europe, reaching Britain by 14,000 years ago, where it is called Creswellian. The exchange of stone and shells over 700 km (435 miles) indicates that social networks were open and covered vast areas. Conical points and harpoons appear for the first time in the Late Magdalenian.
People congregated at certain seasons at very large sites, occupied for several months of the year. These often contain mobiliary art, such as exquisitely carved animal heads, engraved tools such as shaft-straighteners and atlatl crooks, notated bones, and stylized female carvings. Ninety percent of painted and engraved cave art is Magdalenian, mainly known from Franco-Cantabria, but is also is found in Italy, Germany, and Britain. On the Russian Plain, the culture is called Molodovan, with large, semi-permanent dwellings constructed of mammoth bones, with large quantities of mobiliary art. Examples include:
La Madeleine rockshelter (France)
Altamira cave (Spain)
Lascaux cave (France), which display many complex paintings and engravings;
Mezhirich, Mezin, Gontzy, and Yudinovo (Russia)
The extremely cold Older Dryas (17,000-15,000 years ago) was followed by the warmer Bølling and later Allerød phases of the Late Glacial Interstadial (15,000-13,000 years ago), followed by the cold Younger Dryas (13,000-11,500 years ago). As before, cold eras increased regionalization. In southern France and Iberia, the Azilian industry (after Le Mas d'Azil, France) continued many Magdalenian traditions. In the cold north, people adapted by hunting reindeer and increasing production of microlithic toolkits which continued into the Mesolithic.
LATE PLEISTOCENE DISPERSALS: COLONIZATION OF THE AMERICAS
No convincing evidence exists for human colonization of the Americas prior to the last 15,000 years, perhaps because earlier, northern, glacial behavioral adaptations had not yet been invented. These then permitted a last, major dispersal.
Possible Source Populations During glacial periods, lower sea levels where the Bering Strait now lies created the grassland of "Beringia" connecting Siberia to Alaska. It supported mammoth and Upper Paleolithic hunter-gatherers, who crossed it before 10,500 years ago, when Beringia was submerged for the last time. An ice-free corridor between the Laurentide and Cordilleran ice sheets may have served as a way south. Alternatively, the same people may have crossed the 55-km (18-mile) wide Bering Strait by boat, or along Pacific Rim coasts. A third argument, based on similarities in projectile points in western Europe and the USA states that Solutreans made a trans-Atlantic crossing. This is less plausible, as the Solutrean predates the similar American points by at least 5000 years, and such a crossing required unknown technology.
Archaeological, genetic, and linguistic data point to eastern Eurasia, especially northeastern Siberia, as the source population for the Americas, from one or more population waves. The relatively poor archaeology does not provide direct evidence of Paleoindian forebears, but artifacts, lifeways, and human skeletons at some sites suggest connections:
Uptar (Siberia)
the Diuktai culture, Aldan Basin (Siberia)
Kennewick Man and others (United States) show links to Southeast Asia
Linguistic and genetic evidence point to Asian sources with a complex colonization sequence. Three Native American languages groups, Na Dene, Eskimo-Aleut, and Amerind, may represent a "three wave" colonization. Others argue for one event before 20,000 years ago, after which proto-Paleoindian diverged into the three groups. Genetically, Amerindians fall into five haplotypes or clades that cluster in Siberia. However, there is often little correspondence between linguistic and genetic diversity.
Mitochondrial DNA from living Native Americans is rooted in Siberia or Mongolia, where shovel-shaped incisors are found. Surprising genetic similarity is found among people from Alaska to Brazil, indicating that one dispersal event was primary. Migration from Siberia to Chile occurred by 13,000 years ago, thus some believe that initial dispersal must have been at least 20,000 years ago. The Y chromosome data suggest colonization did not precede 18,000 years ago.
The Archeological Evidence for Pre-Clovis Sites Artifacts associated with extinct Pleistocene mammals were found early in the 20th century at Folsom and Clovis, New Mexico, leading to the concept of the "Clovis horizon" of prehistoric hunters. By the 1960s many believed that colonization could not have occurred before 12,000 years ago, creating the "Clovis barrier" while others argue for pre-Clovis populations. Several sites now indicate that a few humans had arrived by 15,500 years ago:
Meadowcroft rockshelter (United States)
Cactus Hill (United States)
Taima-Taima (Venezuela)
Guitarrero and Pikimachay (Peru)
Monte Verde and Cueva del Medio (Chile)
Pedra Furada (Brazil)
At some sites, ages of 17,500, 22,000, and even 30,000-50,000 years ago have been suggested, but scientific methods cannot yet support such early dates.
Occasional forays by small, highly mobile groups of foragers seem to have occurred 2000-3000 years before the Clovis peoples and their contemporaries. These explorations may have ended in local extinction, or were ephemeral and few in number, making them archaeologically invisible.
The Clovis Phenomenon Archaeological visibility improves with the Clovis phenomenon, suggesting either that more humans arrived or that their behavioral strategies became more likely to leave an archaeological record. The Clovis horizon is based on characteristic projectile points and knives, which were pressure flaked and bear a basal flute to facilitate hafting. Behaviorally, Clovis was once viewed as a fast-moving, west-to-east colonizing blitzkrieg of big-game hunters, seeming to coincide with the extinction of several species of large mammal after it arrived 13,500 years ago. In accordance with the three-wave model of colonization, it was imagined to represent the first (Amerind) migration. Research into Clovis behavior since the 1970s has overturned such simplistic notions. For example, many animals that went extinct are never archaeologically associated with Clovis hunting or diet.
Clovis peoples were not "big game hunters," but generalized foragers, not surprising, as the late Pleistocene environment was complex, with a broad variety of resources. Sites yield evidence that plant foods, and the implements for preparing them, were important:
Shawnee Minisink, Pennsylvania (United States)
Blackwater Draw 1, New Mexico (United States)
Clovis populations did hunt mammoth and mastodon, even using them wastefully, but only as an occasional part of a fluid, technologically simple, generalist strategy.
Synchronically with Clovis, generalized foragers were exploiting plant and small animal resources in the Amazon, such as at Pedra Pintada and other sites in Brazil. Generalized lithic technologies, rather than special points, as in Clovis, facilitated plant processing. Similar generalized strategies were used in Mexico by 12,900 years ago, where stable isotope studies show plant protein was important in human diet. On the Argentinean Pampas, humans, alongside environmental change, do seem to have played a role in the extinction of mastodon and horse.
Box Features
Key Controversy: Modern Human Emergence In Asia
Key Controversy: The Evolution Of Language
Key Controversy:
Homo floresiensis: A Small-bodied Hominin from Indonesia
Key Controversy: The Meaning Of "Venus" Figurines
Key Controversy: Kennewick Man
Key Controversy: Big-Game Extinctions in North America
Key Controversy: The Initial Upper Paleolithic and the Emergence of Modern Behavior
Key Site: Klasies River Mouth: Middle Stone Age Hunters?
Key Sites: Four Sites with Upper Paleolithic Art
Key Site: Monte Verde, Chile
Key Method: Radiocarbon Dating
Key words and terms Chapter 4
Climate, geography, environment
unstable climatic change
glacial periods
deep-sea cores
ice cores
saw-tooth curves
geological and faunal indicators of environmental zones
glacial/interglacial
stadial/interstadial
Eurasia: warm/cold
Africa: cycles of arid or wetter periods
Dating methods
biostratigraphy
electron spin resonance (ESR)
uranium series dating
thermoluminescence (TL)
optically stimulated luminescence (OSL)
Species
Homo sapiens
Homo ergaster
Homo erectus
Homo heidelbergensis
Homo neanderthalensis
Hypotheses, theories
multi-regional evolution hypothesis
regional continuity in fossils
predicts ancient coalescence of modern human genes and similar genetic variability across the Old World,
Single origin evolution hypothesis
Noah's Ark hypothesis, Garden of Eden hypothesis, Out of Africa hypotheses.
replacement of older, indigenous forms of Homo by a modern form
predicts that only Africa will contain transitional and the earliest modern fossils and Africa will have the greatest genetic diversity
Other theories
hybridization and replacement model
assimilation hypothesis
multiple dispersals Out of Africa model
People
Franz Weidenreich
Alan Thorne
Milford Wolpoff
Louis Leakey
W. W. Howells
Chris Stringer
Peter Andrews
Gunter Bräuer
Fred Smith
Marta Mirazón Lahr
Robert Foley
Leslie Aiello
Richard Klein
Philip Rightmire
T.F. Dreyer
Karl Butzer
Rebecca Cann
John Relethford
Henry Harpending
Lewis Binford
William King
Jean-Jacques Hublin
Mary Stiner
Sabine Gaudzinski
Steven Kuhn
François Bordes
Paul Mellars
Daniel Lieberman and John Shea
Joseph Greenberg
Christy Turner
Stephen Zegura
Ryk Ward
Thomas Dillehay
Modern Human Anatomy
cranial capacity usually in excess of 1350 cc
vertical frontal bone (forehead)
high and parallel-walled cranial vault
rounded occipital region
lacking the occipital torus
flat basicranial angle
non-continuous brow ridge
flat, non-projecting face
face below the expanded frontal braincase
distinct chin
limb bones long relative to trunk
distal limb segments long relative to the overall limb
long bones less robust than archaic Homo species
Evolutionary process
encephalization
three chronological groupings:
1) earlier than 250,000 years ago, in which
Homo sapiens traits are evident but a number of
Homo ergaster traits are retained
2) a transitional group with variable primitive and modern traits, dating to between 250,000 and 125,000 years ago
3) a group comprising clear anatomically modern humans dating to less than 125,000 years ago
Fossils, proposed species
the Bodo cranium
Homo rhodesiensis
H. helmei
Omo 1 and 2
Homo sapiens idaltu
LH18
Irhoud 1 and 2
Skhul V cranium
Deep Skull
Pestera cu Oase mandible
Lagar Velho Boy
Genetic evidence
nuclear DNA
blood groups
mitochondrial DNA (mtDNA)
Y chromosome
mutation rate
molecular clock
coalescence
recent coalescence
population bottleneck
DNA sequences coalesced in Africa around 200,000 years ago (mtDNA), a little earlier (nuclear DNA), or a little later (Y chromosome).
"Mitochondrial Eve"
rate of mutation problems
microsatellite DNA
emerging consensus: supports hybridization and replacement hypotheses
population genetics
MtDNA diversity
gene flow in vs. Out of Africa
proto-African, proto-European, and proto-Asian groups
Neanderthal mtDNA
genetic dissimilarity between Neanderthals and modern humans
Modern human behavior
after 100,000 years ago
broad coincidence with the biological emergence of anatomically modern Homo sapiens
Middle Stone Age and Late Stone Age sites in Africa
Upper Paleolithic Europe
Artifactual Evidence
pierced shell, bone, eggshell
stone jewelry
bone points
geometric microliths
possible arrowheads
composite tools
blade technology
Stillbay points
Hunting and Dietary Evidence
broad-spectrum economies
fish remains
shellfish
Cape fur seals
plant residues
grinding stones
large mammals
Middle stone age: active and efficient hunting vs. Kill profile possibly indicating incompetent hunting or scavenging.
Site Modification and Art
ocher crayons to create symbols, or art
cross-hatched engravings on ocher
organized use of space not too convincing in Middle Stone Age
possible "walls" of stone cobbles
windbreaks or simple structures
regular lighting of hearths
cut marks, defleshing of crania, mortuary ritual
debate over gradual vs. relatively sudden emergence of fully modern behavior
Colonization of the Old and New worlds by Homo sapiens
Levantine corridor for movement between Africa and Eurasia
Israel by 110,000-90,000 years ago
Levalloiso-Mousterian toolkits
Neanderthals hunted evenly over dry and wet seasons; modern humans only during the dry season: differing exploitation/mobility
strategies; modern humans more efficient
South Asia, Australia, Caucasus, southeast Siberia, Europe by 40,000 years ago
island-hopping water crossings to New Guinea, Australia, Tasmania
Southern, Eastern Europe by 45,000 years ago
Iberia by 40,000 years ago
Upper Paleolithic
Aurignacian technocomplex
Europe 35,000 years ago, northern European plain 30,000 years ago
carinated and nosed-end scrapers
"strangulated" blades
small, retouched bladelets
bone and antler points
flake technology
prismatic blade technology
awls, needles
tailored clothing
art
engraved ivory and bone
painted and engraved cave walls
human and animal figurines in mammoth ivory
Neanderthals
adaptation to cold conditions
physically demanding lifestyle
Anatomy
retained the cranial and postcranial body of
H. heidelbergensis
large brain
long, low vault
powerful biting musculature that modified the face by buttressing
wear on teeth suggests use as a vice-like tool
large nasal aperture and sinuses to warm and moisten air to protect the brain and retain efficient respiration.
large bodies
robust limbs
pronounced muscular insertions
limbs short relative to trunk
retention of body heat
short lifespan
physical trauma
Exploitation of Resources
mortality peaks in infancy (weaning) and in adolescence (childbirth, hunting)
healed upper body fractures
hunting large animals at close range
plant resources
scavenging and hunting strategies
stable isotope analyses
90 percent of protein from meat
hand-thrust spears, wood or with stone points
opportunistic hunting methods
mammoth and woolly rhinoceros drives
later, prime adult bison and aurochs
small animals, shellfish
Tools, artifacts
Mousterian
Levallois flaking techniques; control shape and size of resulting flakes and blades
discoidal method
edge retouch
scrapers, denticulates, notches, points, bifaces (hand axes)
Bordes' five main Mousterian assemblage types: "Mousterian of Acheulian" tradition, Quina, Ferrassie, denticulate Mousterian, typical Mousterian
Bordes: different Neanderthal groups
Binford: different toolkits reflected different tasks
Mellars: some chronological patterning
The "Mousterian debate"
Neanderthal Behavior
skilled workers of antler and bone
pseudomorphs of wooden items, preserved wooden spear
simple hearths for warmth, light, and cooking, burning wood; coal
organization of activities around hearths
paving of wet floors with stones
use of space fairly simple: disposal and messy tasks, eating, resting, and maintenance
some buried dead: at least 20 clear examples
used ocher fragments
engraved fossilized sea urchin
no convincing examples of art
Neanderthal - modern human interaction
Aurignacian overlap with the late Neanderthals
"Transitional" industries
latest Neanderthal presence 30,000 - 28,000 years ago
period of co-existence or interaction
Neanderthals adopt some H.sapiens behaviors
Middle Paleolithic assemblages with true blade technology and Upper Paleolithic tools, animal teeth pierced and grooved for suspension, bone discs
fossils with possible mix of modern and Neanderthal characteristics
interbreeding debate
more than one cause in disappearance of the Neanderthals.
Modern humans in Europe
Aurignacian ends 28,000 years ago.
Gravettian technocomplex: 29,000 - 21,000 years ago
southern and eastern Europe, Epigravettian to c. 14,000 years ago
Last Glacial Maximum
semi-sedentism: semi-subterranean dwellings organized in an oval
pits cut into the permafrost for storage of meat
highly organized campsites
backed blades, burins, end-scrapers: working of skins, wood, bone, and antler
bow and arrow
spearthrower (atlatl)
cordage, basketry, and textiles
simple firing of loess small animal figurines for rituals
broad spectrum diet: fish and other small animals
stable isotope analysis indicate 50% riverine protein
over 40 burials with ornamentation, grave offerings
social complexity
mobiliary art
"Venus" figurines
cave painting and engraving
hand prints, human figurines
climatic minimum, 21,000-18,000 years ago
southern European refuges
Epigravettian
Solutrean
heat-treatment of stone
leaf-shaped points
climatic amelioration from 18,000 years ago; recolonization
Magdalenian technocomplex
Creswellian
Molodovan
open and vast social networks
conical points, biserial and uniserial harpoons
large-scale, long-term site occupation
Franco-Cantabrian mobiliary art, paintings, and engravings
semi-permanent mammoth bone dwellings
highly decorated tools
climatic oscillations
cold stadials: Older and Younger Dryas (17,000-15,000 and 13,000-11,500)
Late Glacial Interstadial (15,000-13,000 years ago Bølling and Allerød phases
increasing regionalization
Azilian industry
Mesolithic
Colonization of the Americas
routes, source populations
Beringia; Bering Strait, linked Siberia to Alaska
submerged by 10,500 years ago
ice-free corridor between the Laurentide and Cordilleran ice sheets
travel by boat, either across the Bering Strait or along Pacific Rim coasts
Western European Solutreans cross the Atlantic
archaeological, genetic, and linguistic data point to eastern Eurasia
similarity between fluted and unfluted points in Siberia and Alaska
Diuktai culture, Siberia
Kennewick Man
Baja California skulls resemble Southeast Asian populations
linguistic evidence shows complex colonization
Eskimo-Aleut, Amerind, Na Dene language groups
three-wave model vs. single event, further back in time
mitochondrial DNA from living Native Americans is rooted in Siberia or Mongolia,
similarity between geographically separate people indicates one main dispersal archaeological evidence suggests 20,000 years ago
recent genetic data from Y chromosome suggest 18,000 years ago
"Clovis horizon;" "Clovis barrier"
sites now break the Clovis barrier
pre-Clovis as early as 15,500 years ago
Clovis: fast-moving, west-to-east colonizing big-game hunters
blitzkrieg model: Clovis caused extinctions
recent: overturn simple models
Clovis were generalized foragers, large and small animal, plants
fluid, technologically simple
Clovis points
mammoth, mastodon
pressure flaking
fluted points
Sites
Bodo (Ethiopia)
Broken Hill/Kabwe (Zambia)
Elandsfontein/Saldanha (South Africa)
Ndutu (Tanzania)
Salé (Morocco)
Florisbad (South Africa)
Omo/Kibish (Ethiopia)
Jebel Irhoud (Morocco)
Ngaloba/Laetoli (Tanzania)
Singa (Sudan)
Eliye Springs (Kenya)
Herto (Ethiopia)
Die Kelders Cave (South Africa)
Equus Cave (South Africa)
Cave of Hearths/Makapansgat (South Africa)
Border Cave (South Africa)
Klasies River Mouth (South Africa)
Dar-es-Soltan (Morocco)
Taforalt, Afalou, and Nazlet Khater (North Africa)
Taramsa (Egypt)
Mugharet es-Skhul (Israel)
Jebel Qafzeh (Israel)
Neanderthal 1 (Germany)
Vindija Cave (Croatia)
Mezmaiskaya Cave (Caucasus)
Blombos Cave (South Africa)
Mumba (Tanzania)
Howieson's Poort (South Africa)
White Paintings Shelter (Botswana)
Herolds Bay caves (South Africa)
Sea Harvest (South Africa)
Hoedjies Punt (South Africa)
Swanscombe (England)
Mauer (Germany)
Sima de los Huesos/Atapuerca (Spain)
Krapina (Croatia)
Kebara Cave (Israel)
Dederiyeh Cave (Syria)
Gorham's Cave (Gibraltar)
Pagnolo d'Asolo (Italy)
Gröbern (Germany)
Umm el Tlel (Syria)
La Cotte de Saint Brelade (Jersey)
Mauran (France)
Wallertheimin (Germany)
Il'skaja (Russia)
Gruta do Caldeiräo (Portugal)
Latium (Italy)
Vanguard Cave (Gibraltar)
Le Moustier (France)
Salzgitter-Lebestedt (Germany)
Buran-Kaya III/Crimea (Ukraine)
Abric Romaní (Spain)
Lehringen (Germany)
Les Canalettes (France)
Shanidar Cave (Iraq)
La Ferrassie rockshelter (France)
Tata (Hungary)
Amud cave (Israel)
Tabun cave (Israel)
Dederiyeh Cave (Syria)
Ksar Akil (Lebanon)
Üçagizli Cave (Turkey)
Niah Cave (Sarawak)
Willandra Lakes (Australia)
Kow Swamp (Australia)
Malakunanja rockshelter (Australia)
Lake Mungo (Australia)
Aurignac rockshelter (France)
Bacho Kiro (Bulgaria)
Temnata (Bulgaria)
Grotta de Fumane (Italy)
El Castillo (Spain)
Hohle Fels cave (Germany)
Vogelherd cave (Germany)
Geissenklösterle cave (Germany)
Chauvet cave (France)
Grotte du Renne (France)
Pestera cu Oase (Romania)
Lagar Velho, (Portugal)
La Gravette (France)
Pavlov (Moravia)
Dolní Vestonice I and II (Moravia)
Kostenki I (Russia)
Sungir' (Russia)
Pech Merle (France)
Solutré (France)
Stellmoor (Germany)
La Madeleine rockshelter (France)
Laugerie Haute (France)
Altamira (Spain)
Lascaux (France)
Mezhirich (Russia)
Mezin (Russia)
Gontzy (Russia)
Yudinovo (Russia)
Le Mas d'Azil (France)
Uptar (Siberia)
Folsom (United States)
Clovis (United States)
Meadowcroft rockshelter (United States)
Cactus Hill (United States)
Taima-Taima (Venezuela)
Guitarrero (Peru)
Pikimachay (Peru)
Monte Verde (Chile)
Cueva del Medio (Chile)
Pedra Furada (Brazil)
Shawnee Minisink (United States)
Blackwater Draw (United States)
Debert (Canada)